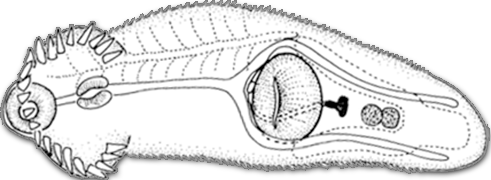
- Parasites, species interactions and food webs
- Bacterial microbiomes of trematodes
- Molecular ecology and diversity of parasites
- Evolution of host specificity
- Transmission strategies of trematodes
- Parasitism, global change and natural ecosystems
- Host manipulation by parasites
- Geographical variation and adaptation in parasitism
- Macroecology and biogeography of parasitism
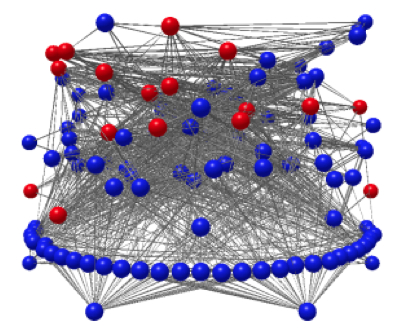
Three-dimensional network representation of the Otago Harbour intertidal mudflat food web. Free-living taxa are shown in blue, and parasitic ones in red. The vertical position of a taxon corresponds to its trophic level.
Parasites, species interactions and food webs: Although traditionally excluded from food web studies, parasites are integral parts of trophic networks. Their presence can modify fundamental interspecific interactions, such as competition and predation. They also contribute to the flow of energy through a food web both as consumers of their hosts, but also as resources themselves (they are eaten by a range of predators). We investigate how parasitism impacts interspecific interactions, and how incorporating parasites into food webs changes their topological structure as well as their stability, resilience and other dynamical properties. Also, we use (i) network analysis tools to determine how food web structure affects parasite transmission, by favouring certain trophic transmission routes over others; and (ii) compound-specific stable isotope analyses to quantify energy flow from host to parasite, and resolve the true nature of host-parasite interactions. Our work focuses on New Zealand lake and coastal ecosystems as model food webs varying with respect to species richness, presence of introduced species, etc.
Bacterial microbiomes of trematodes: Like all other animals, trematodes harbour a microbiome, i.e. a collective of bacteria residing in their tissues. These endosymbionts can alter the phenotype of their trematode host. For instance, if there is a conflict of interest between bacteria and their trematode host, the former could modify the development or transmission strategies of the trematode to their own advantage. Alternatively, when their interests align, the bacteria may cooperate with the trematode and facilitate its host exploitation by contributing novel weapons or abilities. Using metagenomic tools and bioinformatics, we are now characterising the microbiomes of New Zealand trematode species to determine whether endosymbiotic bacteria can shape the phenotypes and strategies of parasites. We are also planning experimental studies of the bacteria-trematode interactions to further explore the impact of microbes on parasites. The project is currently funded by the Marsden Fund.
Molecular ecology and diversity of parasites: Despite the ubiquity and importance of parasites within natural ecosystems, few studies have used genetic data to examine their ecology and diversity. We are currently testing environmental DNA (eDNA) as a tool to detect, quantify and monitor parasite diversity from water samples taken in natural lake ecosystems. On a smaller scale and using trematodes, identifying genetic clones within the same species provides empirical data for addressing questions concerning the number of parasite infections within individual hosts, the influence of environmental conditions on parasite clonal diversity within hosts, the spatial distribution of clones within hosts, trematode transmission strategies, how genetic relatedness affects life history strategies adopted by trematodes, etc. We are investigating several of these questions with New Zealand freshwater and marine trematodes using microsatellite loci to identify genetic clones. In addition, we are also using molecular markers to identify cryptic species and to address large-scale questions of phylogenetics, evolution, phylogeography and gene flow among parasite populations, and making contributions to the taxonomic description of New Zealand's parasite biodiversity.
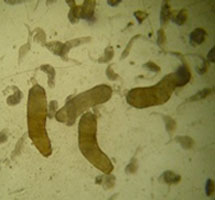

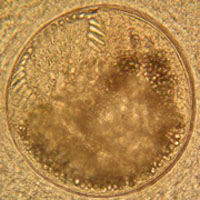
The trematode Acanthoparyphium sp. (Echinostomatidae): rediae and some free cercariae inside the snail first intermediate host (left), free-swimming cercariae (middle), and encysted metacercariae inside a cockle second intermediate host (right).
Evolution of host specificity and host-parasite coevolution: There are constraints on the ability of parasites to expand their host range by exploiting novel host species. We are investigating the role of morphological and genetic variation in determining how parasites perform when exposed to hosts that are progressively more phylogenetically distant from their normal host. The research uses trematodes and other aquatic parasites as model organisms, to explore what affects the ability of parasites to overcome local adaptation and succeed at exploiting geographically distant host populations. We are also looking at the coevolution of host-parasite associations in other sustems, including New Zealand skinks and geckos and their nematode parasites, as well as deep-sea rattail fishes and their helminth parasites, to identify when and under what circumstances parasites switch hosts, and what facilitates parasite sharing among sympatric host species.
Transmission and life history strategies of trematodes: Most parasites show some intraspecific variability in their developmental schedules and transmission pathways. In some trematode species within their first intermediate hosts (snails), we now have evidence that members of the same clone show a division of labour, with some rediae specializing for defence, others for reproduction. In other trematodes within their second intermediate hosts, precocious maturation involving an abbreviated life cycle is commonly observed. Some of this plasticity is under genetic control, but much of it is also influenced by external factors. For instance, the presence of other parasites within the same host can affect the strategies chosen by a parasite: these other parasites can be close genetic kin, unrelated conspecifics, or individuals of other species. Can a parasite adjust its developmental strategy based on the identity of the other parasites with whom it shares the host? Also, parasites in their intermediate host can benefit from information on the current abundance of definitive hosts. For example, trematodes capable of progenesis (early sexual maturation) would benefit by initiating reproduction inside their intermediate host when definitive hosts are rare, thus bypassing the need for transmission to the definitive host to complete the life cycle. Are parasites capable of adjusting their life history strategies based on the likelihood of transmission under current external conditions? We are using field and laboratory experiments, using single and mixed-clone infections of model trematode species in their intermediate hosts, to explore plasticity in development, life history strategies, and transmission routes in parasites.
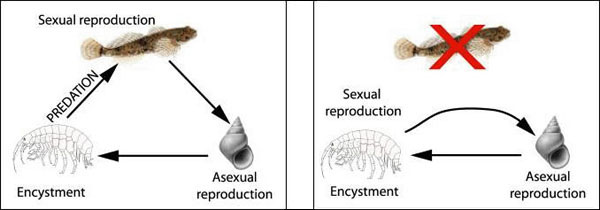
Alternative transmission modes in the trematode Coitocaecum parvum (Opecoelidae): the normal 3-host life cycle (left), and the truncated 2-host cycle in which the parasite reproduces precociously in its second intermediate host (right).
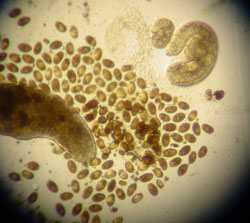
Coitocaecum parvum: progenetic metacercaria (left) with its eggs, and two 'normal' metacercariae (right), including a recently-encysted one. By self-insemination, hermaphroditic worms adopting progenesis can produce several hundred eggs while inside their amphipod intermediate host.
Parasitism, environmental change and community structure in natural ecosystems: Parasites are ubiquitous, though invisible, components of ecosystems. They influence the survival and reproduction of individual hosts, the dynamics of host populations, and the structure of entire communities. In marine systems, our research focuses mostly on parasitism in molluscs and crustaceans. Using field and lab experiments, we study the transmission of parasites in these systems, as well as the direct and indirect impact of parasitism on the density of key host species, the biomass of key functional groups, and the overall biodiversity of intertidal ecosystems. We explore the likely effect of harvesting, species invasion, global warming, ocean acidification, microplastic pollution and other environmental changes on the interaction between parasitism and the structure and diversity of coastal systems. In freshwater systems, we are investigating the impact of parasites on the structure and dynamics of planktonic communities, as well as on entire lake communities. We also study the interaction between parasitism and biological invasions, i.e. when exotic species become hosts to native parasites, and at the impact of trematode parasites on juvenile fish, following up evidence that infection early in life can cause malformations in fish and affect survival and recruitment to the adult population. In terrestrial systems, we are investigating the interactions between avian malaria, bird migration and the composition of bird communities across environmental gradients, taking advantage of the fact that the relative abundance of native and exotic bird species, or migrant versus resident species, changes spatially.


When heavily infected by echinostome trematodes, cockles lose their ability to burrow and simply lie at the sediment surface (left). Echinostome metacercariae accumulate in a cockle's foot until its function is impaired. This has major implications for the benthic community, because surfaced cockles alter habitat properties such as sedimentation rates and availability of hard substrate, with consequences for other organisms. There may also be effects on other parasites living in cockles, such as the copepod Pseudomyicola spinosus (right).

The acanthocephalan Acanthocephalus galaxii is a common intestinal parasite of New Zealand freshwater fishes, but is very rare among amphipod intermediate hosts. Can it alter amphipod behaviour in ways that increase its transmission success to fish?
Manipulation of host behaviour or phenotype by parasites: Numerous taxa of parasites are capable of modifying the appearance or behaviour of their host in ways that increase the probability that the parasites will achieve transmission and complete their life cycle. Larval helminths that must pass from an intermediate host (prey) to a definitive host (predator) via the food chain are notorious for their abilities to enhance their host's susceptibility to predation. Nematomorphs and mermithid nematodes that must reach water from a terrestrial host are also capable of host manipulation. Our current research focuses on the implications of parasite manipulation for animal behaviour in general, by quantifying how manipulation by parasites may influence the entire behavioural repertoire of the host, and looking at whether behavioural strategies evolve in different ways in populations exposed and not exposed to manipulating parasites. We are also investigating the mechanisms underlying altered behaviours in infected hosts, using a range of -omics approaches.

Encapsulated (melanized) metacercariae of Maritrema novaezealandensis in a resistant amphipod. The metacercariae are visible as three reddish dots through the amphipod's exoskeleton; they were killed by the host immune system following penetration.
Geographical variation and local adaptation: Different populations of the same host species experience different levels of infection, and are also exploited by different parasite species. We investigate the factors responsible for these spatial patterns in parasitism. Also, immunity to infection comes at a cost: any energy allocated to fighting parasites is unavailable for other functions, such as reproduction. We thus expect natural selection to optimize investments in immunity such that they reflect actual exposure to parasites: if infection is likely, greater immunity should be favoured, and vice versa. We are comparing the ability of crustaceans from different populations to resist infection by parasites, to determine whether individuals from areas where parasites are absent are more susceptible to infection and less capable of an immune response than those from populations facing high levels of parasitism.
Macroecology and biogeography of parasitism: Large-scale ecological patterns are the outcome of both evolutionary processes and historical biogeographical events. The way in which parasite species within any given taxon are distributed among host species or among geographical areas reflect the action of these forces, and these distribution patterns can shed light on which force has played the most important role in shaping parasitism in natural systems. Using large databases on host-parasite associations, and a range of phylogenetic, biogeographical and macroecological approaches, we search for emergent patterns in parasite ecology. In particular, we have recently investigated global patterns in parasite biodiversity, as well as patterns of parasite specialization in host-parasite networks. The taxa we investigate range from helminths in fish to fleas on mammals. Our findings are casting a new light on the general importance of parasitism as a component of complex ecological systems.