The text below is modified slightly from an original review which appeared as:
Fordyce, R. E. 2006. New light on New Zealand Mesozoic reptiles.
Geological Society of New Zealand newsletter 140: 6-15.
Thanks to the President and Editor of the Geological Society for permission to post this review. You can get a pdf of the article.
Jeff Stilwell and coauthors recently (early 2006) published the first report of dinosaur bones from Chatham Island. The fossils include convincing material, and the occurrence promises more finds. Questions remain, however, about the stratigraphic setting. This commentary summarises the recent finds, considers earlier reports of New Zealand Mesozoic vertebrates, and reviews some broader issues of Mesozoic reptile paleobiology relevant to New Zealand.
The Chatham Island finds
A diverse team reports on the Chathams finds. Jeff Stilwell (fig. 1 here) is an invertebrate paleontologist with research interests on Gondwana breakup and Southern Hemisphere Cretaceous-early Cenozoic molluscs (e.g. Stilwell & Zinsmeister 1992), including Chatham Islands (1997). Several authors are vertebrate paleontologists – Chris Consoli, Tom Rich, Pat Vickers-Rich, Steven Salisbury, and Phil Currie – with diverse experience of dinosaurs. Rupert Sutherland and Graeme Wilson (GNS) are well-known for their research on tectonics and biostratigraphy.

Fig 1 Jeff Stilwell in the field, Seymour Island, Antarctica photo E Fordyce
The Chathams article describes a range of isolated bones attributed to theropod (“beast-footed,” carnivorous) dinosaurs, including a centrum (main part or body of a vertebra), a pedal phalanx (toe bone), the proximal head of a tibia (lower leg bone, at the knee joint), a manual phalanx (finger bone) and a manual ungual (terminal “claw” of a finger). On names of groups, fig. 2 here gives a simple guide to dinosaur classification. Specimens are catalogued in the GNS collections. Those outside the field of vertebrate paleontology might be sceptical that isolated and sometimes incomplete bones can be identified as to position (e.g. vertebra, limb) or to species. However, this approach has been thoroughly tested since Georges Cuvier proposed the idea of “correlation of parts” – that, because organisms can be viewed as integrated whole entities, a species may be identified from only a fragment of a skeleton. In vertebrate paleontology, as in human forensic pathology, the identification of single bones is based on knowledge of more-complete specimens. There is a wealth of supporting information in the global literature (e.g. Weishampel et al. 2004, Farlow & Brett-Surmann 1997), and the technique is widely and successfully used. Of the Chathams bones, the phalanges are compellingly dinosaurian, although it is also possible that the ungual phalanx is from the foot of a large bird. The vertebral centrum does not seem to represent a marine reptile (plesiosaur, mosasaur). For this incomplete element, the authors state that position in the vertebral column and orientation are uncertain, but their conclusion that it was from a medium-sized theropod about 4 m long and 3 m high suggests far more confidence. The fragment of tibia (reportedly proximal, and thus figured upside down) cannot reasonably belong to other than dinosaur.
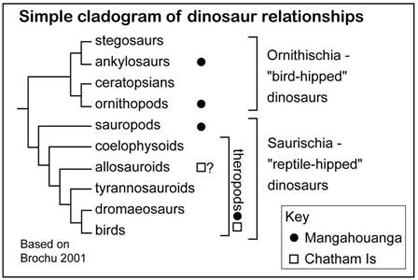
Fig 2 Simple cladogram of dinosaur relationships, based on Brochu 2001). Symbols indicate groups reported from the New Zealand Cretaceous
The stratigraphic setting is intriguing. The bones are from a coastal outcrop of Takatika Grit on the northern part of Chatham Island (see also Campbell et al. 1993: 60). This unit is a thin (~10 m) marine unit, with abundant phosphorite nodules, greensand and volcanogenic debris, resting unconformably on basement schist. Stilwell et al. (2006: fig. 2) divide the unit into 14 beds, of which 8 contain bone (although p 249 states 3 bone horizons). The article title will be taken widely to mean that the KT boundary is present in this sequence, will ensure wide citation, and will likely smooth the path to more research funding. Note, though, that the authors actually state: “The grit may represent, in part, an allochthonous accumulation of reworked uppermost Cretaceous sediments and fossils that were deposited post K–T boundary, in the Danian. However, the mixed latest Cretaceous–earliest Tertiary age of associated microfossils indicate that the K–T boundary may be present below the exposed bone-rich beds…” It's interesting to read “may be present,” given the lack of a firm Haumurian (late Cretaceous) or older age for the grit. Later (p 248), it is stated that the Takatika Grit is significant for allowing reconstruction of paleoenvironment and biota from just before the KT boundary, hinting at a Cretaceous age after all. Readers would have been served better had ages been indicated directly on the stratigraphic column of Stilwell et al.: fig. 2. I await a more-conclusive statement on age.
For now, we can assume that Cretaceous terrestrial bones were reworked into Paleocene shallow marine sediments. Is that really possible? Yes; dinosaur bones and remains of other terrestrial organisms have been collected many times from shallow marine strata. Consider, for example, Late Cretaceous dinosaurs from shallow marine Tahora Formation of Mangahouanga Stream, Hawkes Bay (Molnar and Wiffen 1994), and Late Eocene terrestrial mammal bones from shallow marine La Meseta Formation of Seymour Island, Antarctica (Reguero et al. 2002).
As with many good stories, there is an earlier history. Campbell et al. (1993: 62-64) mentioned the presence of reptile bones, possibly dinosaurian (see their fig. 4.30), in the Takatika Grit, but didn't offer a firm identification. Before that, in 1991, University of Canterbury Geosciences student Evan Meek reported bone associated with phosphorite horizons in the Grit, and collected several fragments of presumed plesiosaur bone now held in the Geology Museum at University of Otago.
Some points made by Stilwell et al. don't stand close scrutiny. For example, it is implied twice in the introduction to the paper that the dinosaurs are from an oceanic island, and on p 249 the authors emphasise that the dinosaurs are from an isolated setting nearly 1000 km east of New Zealand. These statements miss the point. “Oceanic” is correct in the sense that Chatham Island is now distant from mainland New Zealand, but in the geological sense and especially in the paleobiogeographic sense it is misleading, and the authors should know better: Chatham Island is based on New Zealand continental crust, not oceanic crust. Some dinosaur afficionados likely to read Stilwell et al. without knowledge of New Zealand geology could be confused needlessly by such wording. Stilwell et al. (their fig. 4) indeed show Chatham Island as continuous with what is now South Island, as also indicated in Fig. 3 of this review (based in turn on reconstructions by King).
Mesozoic terrestrial reptiles
The article on the Chathams dinosaurs is one of many on Mesozoic, mainly Cretaceous, reptiles from New Zealand. Our record of marine reptiles is highly informative, yet the more fragmentary record of dinosaurs – perhaps a shoe-box full of bones in contrast to a shipping-container full of marine reptiles – arouses interest because of implications to ecology and paleogeography. Early geologists hinted inconclusively at the presence of terrestrial reptiles. For example, Haast (1870: 189) commented, about Cretaceous reptiles at Waipara, “There was also, what I consider to be, the distal or lower portion of the femur, which, judging from the articulation, evidently had some affinities with terrestrial remains, such as the Iguanodon a herbivorous terrestrial reptile, of enormous size, and living in the Northern Hemisphere, in the wealden and greensand (Cretaceous) period” [original spellings and capitals retained]. Later, an anonymous author, most likely Hector (in McKay 1877: 41), referred to the tooth of a "megalosauroid . . . land saurian", apparently also from Waipara. The identity of these specimens was not elaborated on and, by 1900, there was still no firm evidence of dinosaurs in New Zealand. In following decades, many people assumed that those reptiles had never actually been here, and the absence of evidence was taken as evidence of absence. Fleming (1962: 93), however, cautioned in generalising about the absence of fossil land vertebrates, particularly as some living forms, such as tuatara, must have had a long Tertiary record in spite of a lack of fossils.
As it happens, in 1958, Don Haw had already discovered reptile bones in Cretaceous shallow marine strata of Mangahouanga Stream, Hawkes Bay (Haw 2002), stimulating the now widely-recognised field programme of Joan and Pont Wiffen and associates including Crabtree, McKee, and Moisley. The first formal report of a New Zealand dinosaur was by Ralph Molnar (1981), who described an isolated vertebra recovered by the Wiffen group from Mangahouanga. The bone did not match any of the marine reptiles already known from New Zealand. Molnar, a Brisbane-based dinosaur paleontologist trained in USA, convincingly argued that the vertebra recovered by Wiffen was from the tail of a theropod dinosaur. Later finds, reviewed by Molnar and Wiffen (1994) and Wiffen (1996), include fragments of other dinosaurs: the partial rib from a sauropod (large quadrupedal plant-eating saurischian), the pelvis of a hypsilophodontid (an small bipedal ornithopod), a fragment of ankylosaur (armoured ornithopod), and the phalanx of a probable theropod (but possibly a large bird). Reptiles other than dinosaurs are known from Mangahouanga, including a protostegid marine turtle (Wiffen 1981), and a fragment of pterosaur (Wiffen and Molnar 1988). Beyond the primary literature, these discoveries are well-covered in popular books (e.g. Stace and Eagle 2001).
What of New Zealand localities other than Chatham Island and Mangahouanga Stream? The only other described putative dinosaur is an isolated manual phalanx (finger bone) from the Huriwai Measures (Puraroan, Late Jurassic), south of the Waikato River mouth. This single small bone was identified as probable theropod (Molnar et al. 1998); it represents the first tetrapod vertebrate described from the New Zealand Jurassic.
There are hints of other tetrapod fossils from our Mesozoic, although these are not necessarily dinosaurian and some may actually represent marine groups. Fleming et al. (1971) mentioned the "teeth having Labyrinthodont characters" earlier reported by Hector (1880, 1886) and Worley (1894) from unspecified formations (probably Triassic) at Nugget Point, South Otago, and Wairoa district, Nelson. These could represent ichthyosaurs or, alternatively, an amphibian. Fordyce et al. (2003) recently reported a bone fragment from a large “labyrinthodont” amphibian (strictly, a sterospondyl) from the basal Triassic south of Mataura.
Mesozoic marine reptiles
The New Zealand record of marine reptiles, particularly Piripauan-Haumurian (Late Cretaceous), is significant at a global level, thanks to some rather complete and informative specimens. The key groups are plesiosaurs and mosasaurs, while the main localities are Mangahouanga Stream, Haumuri Bluff-Conway, Waipara, and Shag Point. For a historical overview, and details of some of the important species, one starts with the monograph of Welles & Gregg (1971). That document arose from Welles' visit to New Zealand in the late 1960s (Gregg 1997) to start a new phase of study on Cretaceous reptiles after a break of about 70 years since the days of Hutton. Welles and Gregg reviewed the early contributions of Owen, Haast, Hector (especially 1874), and Hutton, and the productive field work of the indefatigable Alexander McKay. The monograph also gave details of unpublished early research, and discussed the tragic loss of fossils collected by McKay and sent overseas for study. A notable contribution of Welles and Gregg was to name and describe a new species of mosasaur (sea-lizard), Prognathodon waiparaensis, based on a partial skull recovered from just below the KT boundary in the Waipara River section at Laidmore-Claremont. The taxonomy in the Welles and Gregg monograph is specialised, and when I read the article as a graduate student seeking an introduction to reptile anatomy and classification, I found it not very helpful; other sources, such as books by A.S. Romer, were more rewarding. (For more recent works, see Callaway & Nicolls 1997, and many articles in Journal of Vertebrate Paleontology.)
Much research on marine reptiles has been reported since 1971, and only a brief summary is given here. Material from Mangahouanga Stream (Wiffen and Moisley 1986) includes the new plesiosaur Tuarangisaurus and more material attributed to Mauisaurus, plus mosasaurs (Wiffen 1981a, 1990) including a Prognathodon and 3 new species: Moanasaurus mangahouangae, Rikisaurus tehoensis, and Mosasaurus flemingi. It would be surprising to find a diversity of closely related species (in this case, mosasaurs) of similar size in one assemblage, and whether all the latter mosasaurs are distinct is moot.
To consider fossils from northern South Island, Caldwell et al. (2005) reported new material of the mosasaur Taniwhasaurus oweni (including T. haumuriensis of previous authors) from Haumuri Bluff. Norton Hiller and Al Mannering at Canterbury Museum have made notable advances. Hiller et al. (2005) described rather complete and highly informative new material of the plesiosaur Mauisaurus haasti from Ngaroma, Conway River, clarifying the concept of a species named by James Hector, and confirming M. haasti as an elasmosaur (extreme long-necked plesiosaur). Because Tuarangisaurus keyesi and Mauisaurus haasti are not known from comparable material (the first is based on a skull, the latter on postcranial skeleton), it is not clear whether the 2 species are truly distinct. A possible third elasmosaur species may be represented by an indeterminate fragmentary elasmosaur (Hiller & Mannering 2004) from the classical locality of Claremont-Laidmore at Waipara. Yet more material is known from Waipara: most recently, Hiller & Mannering (2005) described a cluster of postcranial remains from a large concretion, apparently representing another elasmosaur distinct from Mauisaurus haasti. The species was not named because it lacks a skull – probably the single most important element on which to base a new species. The most-recently recovered Waipara reptile is a mosasaur, apparently a new Prognathodon, which includes the skull and jaws. The fossil is under preparation (see www.otago.ac.nz/geology/features/paleontology/).
From the south, the large plesiosaur Kaiwhekea katiki (Shag Point) was described by Cruickshank & Fordyce (2002) as a new genus in the mainly Jurassic family Cryptoclididae - a group in which the neck is significantly shorter than in elasmosaurs. Kaiwhekea is notable as (currently) being the only New Zealand Mesozoic reptile represented by an articulated and nearly complete skeleton, albeit with many bones represented by natural moulds. Kaiwhekea was compared with 2 other Gondwanan plesiosaurs, Aristonectes and Morturneria, also thought to be cryptoclidids. Later, Gasparini et al. (2003) showed that Aristonectes is probably a specialised elasmosaur. Where does Kaiwhekea belong? It is not an elasmosaur in the usual sense of the term. O'Keefe (2004) used cladistic analysis to show that Kaiwhekea might not belong in the Cryptoclididae, but rather in the specialised cryptocleidoid family Cimoliasauridae. It is trite but true to say that more research is needed.
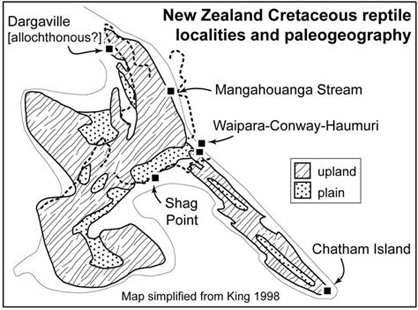
Figure 3. New Zealand Cretaceous reptile localities and paleogeography, based on Cruickshank and Fordyce (2002) with map simplified from King (1998).
Beyond mosasaurs and plesiosaurs, a third marine reptile group is known from the New Zealand Mesozoic, the dolphin-like Ichthyosauria. Fleming et al. (1971) reviewed the specimens, most of which were reported first in the 19th century. Probably the most tantalising fossil known by 1971 was the Late Triassic partial rostrum reported by Campbell (1965) from the Murihiku Supergroup. More Murihiku specimens have been found since, but they are too incomplete to identify (Fordyce 2003). Sachs & Grant-Mackie (2003) recently reported a fragment of snout of a presumed Cretaceous (and thus geologically young) ichthyosaur from Northland, giving hope of finding other more-complete material.
Mesozoic New Zealand – isolated in a high latitude setting
For the Jurassic and much of the Cretaceous, New Zealand was probably within or close to the polar circle (e.g. Lawver et al. 1992, Pole 1999, McLoughlin 2001, Schellart et al. 2006), implying significant seasonal variation in light and climate, and thus in biological productivity (plant growth). Further, by about 85 Ma (Piripauan; Cooper 2004), the Tasman Sea was opening, and the New Zealand landmass was presumably physically isolated from other parts of Gondwana. There are implications for dinosaur (and marine reptile) biology, as noted by Molnar and Wiffen (1994) for New Zealand, and by Rich et al. (2002) for southern Australia. In summary, in a high latitude setting, dinosaurs on an “island continent” would have had limited ability to migrate during winter cold and darkness. Marine reptiles would likely be affected by fluctuations in food caused by great seasonal variation in productivity. How many dinosaurs lived on Late Cretaceous New Zealand after separation from the Gondwana margin? Molnar (1981) noted that a small landmass would support only a small population of any one dinosaur species, and citing small population numbers that might barely be self-sustaining. His discussion, however, did not consider that home ranges of many organisms are not exclusive but may overlap; there may have been more dinosaurs on New Zealand than suggested by Molnar. It is intriguing also that some reptiles may represent late occurrences of groups otherwise reported earlier in the Northern Hemisphere fossils. Consider, for example, the Late Cretaceous plesiosaur Kaiwhekea, which apparently represents an otherwise mainly Jurassic lineage known from Europe, or the Early Cretaceous report of an Allosaurus-like dinosaur, presumably close to Jurassic forms from North America. Another “out of time” record is that of a supposed ceratopsian dinosaur from Victoria, which is older than the start of the great ceratopsian radiation of the northern Hemisphere. (Ceratopsians were “horned” herbivores, such as Triceratops.) Admittedly, those specimens known from nearly complete skeletons (Kaiwhekea) are more convincing than those based on single bones (the Australian Allosaurus and ceratopsian). Some of these issues were discussed further by Long (1998) and Rich and Rich (2000).
References
- Brochu, C. A. 2001. Progress and future directions in archosaur phylogenetics. Journal of Paleontology 75 (6): 1185-1201.
- Caldwell, M. W., Holmes, R., Bell, G. L., & Wiffen, J. 2005. An unusual tylosaurine mosasaur from New Zealand: A new skull of Taniwhasaurus oweni (lower Haumurian; upper Cretaceous). Journal of Vertebrate Paleontology 25 (2): 393-401.
- Callaway, J. M. & Nicholls, E. L. (eds) 1997. Ancient Marine Reptiles. Academic Press, San Diego. 448 p.
- Campbell, H. J., Andrews, P. B., Beu, A. G., Maxwell, P. A., Edwards, A. R., Laird, M. G., Hornibrook, N. de B., Mildenhall, D. C., Watters, W. A., Buckeridge, J. S., Lee, D. E., Strong, C. P., Wilson, G. J., & Hayward, B. W. 1993. Cretaceous-Cenozoic geology and biostratigraphy of the Chatham Islands, New Zealand. Institute of Geological and Nuclear Sciences monograph 2: 1-269.
- Campbell, J. D. 1965. New Zealand Triassic saurians. NZ journal of geology and geophysics 8 (3): 505-509.
- Cooper, R. A. (ed.) 2004. The New Zealand geological timescale. Institute of Geological and Nuclear Sciences monograph 22: 1-284.
- Cruickshank, A. R. I. & Fordyce, R. E. 2002. A new marine reptile (Sauropterygia) from New Zealand: further evidence for a Late Cretaceous austral radiation of cryptoclidid plesiosaurs. Palaeontology 45 (3): 557-575.
- Farlow, J. O. & Brett-Surman, M. K. 1997. (eds), The complete dinosaur. Indiana University Press, Bloomington. 752 p.
- Fleming, C. A. 1962. New Zealand biogeography. A paleontologist's approach. Tuatara 10: 53-108.
- Fleming, C. A., Gregg, D. R., & Welles, S. P. 1971. New Zealand ichthyosaurs - a summary, including new records from the Cretaceous. NZ journal of geology and geophysics 14: 734-741.
- Fordyce, R. E. 2003. Fossils and the history of life. Pp 35-64 in Darby, J. T., Fordyce, R. E., Mark, A. F., Probert, P. K., & Townsend, C. R. (eds), The natural history of southern New Zealand. University of Otago Press, Dunedin. 387 p.
- Fordyce, R. E., Campbell, H. J., and Warren, A. 2003. A presumed stereospondyl (Amphibia, Stereospondyli) from the marine Triassic of Titiroa Stream, Mataura Valley, Southland, New Zealand. Journal of the Royal Society of NZ 33 (1): 301-306.
- Gasparini, Z., et al. 2003. The elasmosaurid plesiosaur Aristonectes Cabrera from the latest Cretaceous of South America and Antarctica. Journal of Vertebrate Paleontology 23 (1): 104-115.
- Gregg, D. R. 1997. Sam Welles in New Zealand. Geological Society of New Zealand newsletter 114: 95-97.
- Haast, J., 1870. Notes on a collection of saurian remains from the Waipara River, Canterbury, in the possession of J.H. Cockburn Hood, Esq. Transactions and proceedings of the NZ Institute 2: 186-189.
- Haw, D. 2002. The discovery of fossils in Mangahouanga Stream. Geological Society of New Zealand newsletter 129: 13-14.
- Hector, J. 1874. On the fossil Reptilia of New Zealand. Transactions of the NZ Institute 6: 333-358.
- Hiller, N. & Mannering, A. A. 2004. Elasmosaur (Reptilia: Plesiosauria) skull remains from the Upper Cretaceous of North Canterbury, New Zealand. Records of the Canterbury Museum 18: 1-7.
- Hiller, N. & Mannering, A. A. 2005. An unusual new elasmosaurid plesiosaur (Sauropterygia) from the Upper Haumurian (Maastrichtian) of the South Island, New Zealand. Memoirs of the Queensland Museum 51 (1): 27-37.
- Hiller, N., Mannering, A. A., Jones, C. M., & Cruickshank, A. R. I. 2005. The nature of Mauisaurus haasti Hector, 1874 (Reptilia: Plesiosauria). Journal of Vertebrate Paleontology 25 (3): 588-601.
- King, P. R. 1998. Paleogeographic reconstructions of New Zealand. Geological Society of NZ miscellaneous publication [Geology and genes symposium] 97: 45-48.
- Lawver, L. A., Gahagan, L. M., & Coffin, M. F. 1992. The development of paleoseaways around Antarctica. American Geophysical Union, Antarctic research series 56: 7-30.
- Long, J. A. 1998. Dinosaurs of Australia and New Zealand, and other animals of the Mesozoic Era. University of New South Wales, Sydney. 188 p.
- McKay, A., 1877. Oamaru and Waitaki districts. NZ Geological Survey report of geological explorations 1876-77, (10): 41-66.
- McKee, J. W. A. & Wiffen, J. 1998. Mangahouanga Stream: New Zealand's Cretaceous dinosaur and marine reptile site [field guide]. Geological Society of NZ miscellaneous publication 96: 1-18.
- McLoughlin, S. 2001. The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Australian Journal of Botany 49 (3): 271-300.
- Meek, E. P. 1991. Sedimentology of two formations within the Tioriori Group, Chatham Island. BSc Hons thesis, Department of Geological Sciences, University of Canterbury, Christchurch. 86 p.
- Molnar, R. E. 1981. A dinosaur from New Zealand. Pages 91-96 in Cresswell, M. and Vella P. (eds), Fifth International Gondwana Symposium, Wellington, New Zealand, 11-16 February 1980. Balkema, Rotterdam.
- Molnar, R. E. & Wiffen, J. 1994. A Late Cretaceous polar dinosaur fauna from New Zealand. Cretaceous Research 15 (6): 689-706.
- Molnar, R. E., Wiffen, J., and Hayes, B. 1998. A probable theropod bone from the latest Jurassic of New Zealand. NZ Journal of Geology and Geophysics 41 (2): 145-148.
O'Keefe, E. R. 2004. On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis, Cope, from Alabama. Journal of Vertebrate Paleontology 24 (2): 326-340. - Pole, M. S. 1999. Structure of a near-polar latitude forest from the New Zealand Jurassic. Palaeogeography Palaeoclimatology Palaeoecology 147 (1-2): 121-139.
- Reguero, M. A., Marenssi, S. A., & Santillana, S. N. 2002. Antarctic Peninsula and South America (Patagonia) Paleogene terrestrial faunas and environments: biogeographic relationships. Palaeogeography Palaeoclimatology Palaeoecology (3-4): 189-210.
- Rich, T. H. V. and Rich, P. V. 2000. Dinosaurs of darkness. Indiana University Press, Bloomington. 222 p.
- Rich, T. H., Vickers-Rich, P., & Gangloff, R. A. 2002. Polar dinosaurs. Science 295 (5557): 979-980.
- Sachs, S. & Grant-Mackie, J. A. 2003. An ichthyosaur fragment from the Cretaceous of Northland, New Zealand. Journal of the Royal Society of NZ 33 (1): 307-314.
- Schellart, W. P., Lister, G. S., and Toy, V. G. 2006. A Late Cretaceous and Cenozoic reconstruction of the Southwest Pacific region: tectonics controlled by subduction and slab rollback processes. Earth-Science Reviews 76: 191-233.
- Stace, G. & Eagle, M. K. 2001. Yes! We had dinosaurs. Penguin, Auckland. 80 p.
- Stilwell, J. D. 1997. Tectonic and palaeobiogeographic significance of the Chatham Islands, South Pacific, Late Cretaceous fauna. Palaeogeography Palaeoclimatology Palaeoecology 136 (1-4): 97-119.
- Stilwell, J. D. & Zinsmeister, W. J. 1992. Molluscan systematics and biostratigraphy of the Lower Tertiary La Meseta Formation, Seymour Island, Antarctic Peninsula. American Geophysical Union, Antarctic Research Series 55: 1-192.
- Stilwell, J. D., Consoli, C. P., Sutherland, R., Salisbury, S., Rich, T. H., Vickers-Rich, P. A., Currie, P. J., & Wilson, G. J. 2006. Dinosaur sanctuary on the Chatham Islands, Southwest Pacific: first record of theropods from the K-T boundary Takatika Grit. Palaeogeography Palaeoclimatology Palaeoecology 230 (3-4): 243-250.
- Weishampel, D. B., Dodson, P., & Osmólska, H. 2004. (eds), The Dinosauria. University of California Press, Berkeley. 861 p.
- Welles, S. P. & Gregg, D. R. 1971. Late Cretaceous marine reptiles of New Zealand. Records of the Canterbury Museum 9: 1-111.
- Wiffen, J. 1981a. Moanasaurus, a new genus of marine reptile (Family Mosasauridae) from the Upper Cretaceous of North Island, New Zealand. NZ journal of geology and geophysics 23: 507-528.
- Wiffen, J. 1981b. The first Late Cretaceous turtles from New Zealand. NZ journal of geology and geophysics 24 (2): 292-300.
- Wiffen, J. 1990. New mosasaurs (Reptilia: Family Mosasauridae) from the Upper Cretaceous of North Island, New Zealand. NZ journal of geology and geophysics 33: 67-85.
- Wiffen, J. 1996. Dinosaurian palaeobiology: a New Zealand perspective. Memoirs of the Queensland Museum 39 (3): 725-731.
- Wiffen, J. & Moisley, W. L. 1986. Late Cretaceous reptiles (families Elasmosauridae, Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand. New Zealand journal of geology and geophysics 29: 205-252.
- Wiffen, J. & Molnar, R. E. 1988. First pterosaur from New Zealand. Alcheringa 12: 53-59.
- Wilson, G. J., Schiøler, P., Hiller, N., & Jones, C. M. 2005. Age and provenance of Cretaceous marine reptiles from the South Island and Chatham Islands, New Zealand. NZ Journal of Geology and Geophysics 48 (2): 377-387.

The author - R.Ewan Fordyce, Associate Professor at Otago University with the jaw of a fossil mosasaur from the Waipara River area
- Introduction
- Otago History
-
Reptiles
- Dinosaurs, plesiosaurs, mosasaurs and more
- Mosasaurs - Prognathodon
- Plesiosaurs - Kaiwhekea katiki
- Central Otago Miocene crocodile
- Dolphins
- Sharks
- Whales
- Fossil penguins
-
Amphibians
-
Geological settings
- Vanished World Trail
- Geology Museum